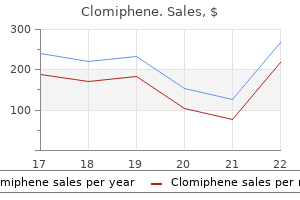
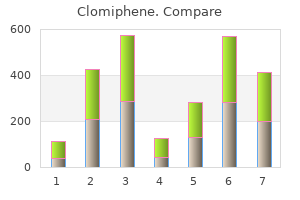
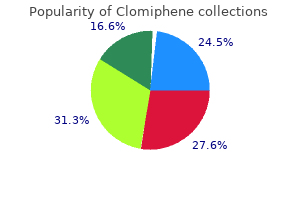
Professor, Department of Obstetrics and Gynecology
University of California at Irvine
Irvine, California
In abduction to approximately 23� menopause 12 months order clomiphene 100mg without prescription, they become nearly purely an elevator and depressor pregnancy vaccines buy cheap clomiphene 25mg, respectively triple negative breast cancer order 50 mg clomiphene fast delivery, of the visual axis pregnancy ovulation calculator discount clomiphene online master card. Superior indirect elevates the posterior aspect of the eyeball women's health oregon city best purchase clomiphene, and inferior indirect depresses it menstruation tracker generic 25mg clomiphene otc, which implies that the previous rotates the visual axis downwards and the latter rotates it upwards, and both actions happen around the horizontal axis. When the attention is within the primary position, the obliquity of each muscles means that they pull in a course posterior to the vertical axis and both therefore rotate the eye laterally round this axis. With regard to the anteroposterior axis, in isolation, superior indirect intorts the eye and inferior oblique extorts it. Like the superior and inferior recti, due to this fact, the two obliques have a common turning motion across the vertical axis but are opposed forces in respect of the opposite two. Acting in live performance, they could therefore assist the lateral rectus in abducting the visual axis, as in divergence of the eyes in transferring consideration from near to far. Again, like the superior and inferior recti, the instructions of traction of the oblique muscular tissues also differ with ocular place, such that they turn into more practically a pure elevator and a depressor as the attention is adducted. Several forms of eye movement are required to ensure that these conditions are met. Moreover, the movements of both eyes have to be near perfectly matched to obtain the advantages of binocularity. Alternatively, they may be grouped into those movements that shift gaze as visual interest modifications, and people who stabilize gaze by sustaining a steady picture on the retina. They have distinct characteristics, and are generated by completely different neural mechanisms in response to different stimuli, but share a common last motor pathway. Movements that shift or stabilize gaze embody saccades, vergence, pursuit and vestibular-generated reflexes (Leigh and Zee 2006). Thus, each visible axes are endlessly and rapidly transferred to new points of curiosity in any a part of the visual field. Binocular gaze is regularly made to travel routes of probably the most variable complexity in analyzing objects of interest within the subject, and each visible axes must be maintained with adequate accuracy to keep away from diplopia. When interest changes to another characteristic of the visual scene, the eyes execute a fast or saccadic motion to take up fixation. If the required rotation is small, the saccade is correct, whereas small supplementary corrective saccades are needed if the shift is substantial. As an example of the latter, in studying a line of print, the eyes make three or four jerky saccades rather than following the road smoothly; the line is usefully imaged only when the attention is stationary, which means that little of the line is seen by the centre of the fovea. In common, response instances and actions are measured in microseconds, amplitude varies from seconds of arc to many levels, with an accuracy of zero. The speed of saccades is assured by an preliminary, barely excessive, contraction of the suitable muscle tissue to overcome orbital viscous resistance. The essential deceleration when the target is fixated is essentially dependent on the elasticity of the extraocular muscles and orbital delicate tissues, and not on antagonistic muscular exercise. A, the connection between the orbital and ocular axes, with the eyes within the main position, where the visual axes are parallel. B and C, the ocular globe in anterior and posterior views to show conventional geometry. D, the orbits from above, showing the medial and lateral recti and the superior rectus (left) and the inferior rectus (right), indicating turning moments primarily across the vertical axis. E, the superior (left) and inferior (right) indirect muscle tissue showing turning moments primarily across the vertical and likewise anteroposterior axes. F, A lateral view to show the actions of the superior and inferior recti around the horizontal axis. G, A lateral view to present the action of the superior and inferior indirect muscles across the anteroposterior axis. H, An anterior view to present the medial rotational movement of the superior and inferior recti across the vertical axis. I, An anterior view to show the torsional results of the superior oblique (intorsion) and inferior indirect (extorsion) around the anteroposterior axis, as indicated by the small arrows on the cornea. The otoliths also reply to the pull of gravity, generating static vestibulo-ocular reflexes associated with head tilt. Because the foveal image is unaffected by torsional movements, the subject is unaware of any visual penalty. The optokinetic response is another visually mediated reflex that stabilizes retinal imagery when a visible scene is rotated a few stationary subject. As the scene modifications, the eyes observe and maintain the retinal picture regular till the eyes shift rapidly in the other way to another area of the visual scene. The full field of regard, quite than small objects inside it, is the stimulus, and the alternating gradual and quick phases of motion that are generated describe optokinetic nystagmus. This reflex capabilities in collaboration with the rotational vestibulo-ocular reflex. In sustained rotations of the physique, the vestibuloocular reflex fades due to the mechanical preparations of the semicircular canals. In darkness, the reflex, which is initially compensatory, loses velocity, and after roughly 45 seconds the eyeballs turn out to be stationary. Because the reflex is already initiated, the relative delay of visual enter is overcome. The integration of the 2 methods is served by an adjunct visible system projection to the vestibular nuclei by way of the inferior olive and cerebellum. The usual method of evoking optokinetic nystagmus in the laboratory or clinic is to current a horizontally shifting pattern of vertical black-on-white stripes whereas the top of the topic is held stationary. Clockwise and anticlockwise curved arrows characterize intorsion and extorsion, respectively. The eyes converge in path of the midline between the two eyes to obtain imagery of the object on each foveas. In addition, the pupils constrict and the eyes accommodate to achieve sharp, centered pictures. Pursuit eye actions are used to monitor a shifting object of visible curiosity, sustaining the image approximately on the fovea. If the angular shift required to monitor the moving object is massive or the object is moving swiftly, the preliminary saccade is regularly inaccurate and a quantity of small corrective saccades are made earlier than monitoring begins. Because the stimulus is visual, the pursuit system response is topic to a comparatively lengthy latency (approximately a hundred msec); the limitation in performance this imposes could additionally be offset by a predictive capacity when object motion follows a regular pattern, and the attention actions modify in anticipation to pace and direction. The vestibular equipment induces a variety of reflex eye movements to compensate for the possibly disruptive results on imaginative and prescient brought on by head and body motion (Ch. Receptors within the semicircular canals reply to active or passive rotational (angular) accelerations of the top. When the physique makes substantial rotational movements, a vestibulo-ocular reflex generates a cycle of responses involving each the shifting and stabilizing of gaze. Body rotation is matched by counterrotation of the eyes so that gaze path is unaltered and clear vision is maintained. Physical constraint limits the rotation to 30� or less and is adopted by a fast saccadic movement of the eyes to another object in the visible scene and the cycle is repeated. Vision is due to this fact clear all through many of the cycle whereas the image is stationary, however at the value of no helpful imaginative and prescient through the transient durations of the saccades. Other reflexes generated by the vestibular system, which induce compensatory eye movements to stabilize gaze, are activated during transient head actions. When the head is sharply rotated in any path, the eyeball rotates by an equal quantity in the wrong way in response to the stimulation of semicircular canal cristae (angular acceleration), and gaze is undisturbed. Brief rotational actions are generally combined with translational actions (linear acceleration) which may be monitored by otolith organs. For example, a linear displacement occurs in strolling as the pinnacle bobs vertically with each stride, and a rotational displacement occurs as the top rolls, invoking otolith and canal responses, respectively, to stabilize the retinal image. The common factor for each type of horizontal gaze movements is the abducens nucleus. It accommodates motor neurones that innervate the ipsilateral lateral rectus and interneurones that project by way of the medial longitudinal fasciculus to the contralateral oculomotor nucleus, which controls medial rectus. A lesion of the abducens nucleus leads to a total lack of ipsilateral horizontal conjugate gaze. A lesion of the medial longitudinal fasciculus produces slowed or absent adduction of the ipsilateral eye, normally related to jerky actions (nystagmus) of the abducting eye, a syndrome known as internuclear ophthalmoplegia (Leigh and Zee 2006). The gaze motor command includes specialised areas of the reticular formation of the brainstem, which receive a big selection of supranuclear inputs. The main region for the technology of horizontal saccades is the paramedian pontine reticular formation, situated on both sides of the midline in the central paramedian a half of the tegmentum, and increasing from the pontomedullary junction to the pontopeduncular junction. Excitatory burst cells make monosynaptic connections with the ipsilateral abducens nucleus. Pause neurones, situated in a midline caudal pontine nucleus, known as the nucleus raphe interpositus, discharge tonically during fixation however cease firing instantly previous to a saccade. They appear to exert an inhibitory affect on the burst neurones and act as a switch to change from fixation to saccadic mode (Ramat et al 2007). The tonic activity of neurones in the nucleus prepositus hypoglossi and medial vestibular nucleus is assumed to provide a watch position sign to maintain the eccentric position of the attention against the viscoelastic forces in the orbit. Vestibular nuclei and the perihypoglossal complex project on to the abducens nuclei. The final common pathway for vertical gaze actions is formed by the oculomotor and trochlear nuclei. The rostral interstitial nucleus of the medial longitudinal fasciculus accommodates excitatory burst neurones that discharge in relation to up-and-down vertical saccadic movements and project to motor neurones concerned in vertical gaze. Neurones in and across the interstitial nucleus of Cajal, which lies barely caudal to the rostral interstitial nucleus of the medial longitudinal fasciculus, provide signals for vertical gaze holding. Lesions throughout the posterior commissure predominantly give rise to disturbances in upgaze, related to different indicators of dorsal midbrain syndrome. Discrete lesions positioned extra ventrally within the region of the rostral interstitial nucleus of the medial longitudinal fasciculus may cause mixed up-and-down, or mainly downgaze disturbances (Leigh and Zee 2006). The vestibulocerebellum (flocculus and nodule) is involved in gaze holding, clean pursuit and the vestibuloocular reflex. The dorsal vermis and fastigial nucleus play a serious role in programming correct saccades and easy pursuit. The cerebral hemispheres are extremely essential for the programming and coordination of each saccadic and pursuit conjugate eye movements (Ch. There appear to be 4 primary cortical areas in the cerebral hemispheres involved in the generation of saccades. These are the frontal eye field, which is positioned on the intersection of the superior precentral sulcus with the superior frontal sulcus (Brodmann space 8) (Amiez and Petrides 2009); the supplementary eye field, which lies within the dorsomedial frontal cortex (Brodmann area 6); the dorsolateral prefrontal cortex, which lies anterior to the frontal eye field in the second frontal gyrus (Brodmann area 46); and a posterior eye area, which lies within the parietal lobe within the medial wall of the posterior half of the intraparietal sulcus, including parts of the supramarginal and angular gyri (Brodmann areas 39 and 40), and the adjacent lateral intraparietal sulcus (M�ri et al 1996). These areas all appear to be interconnected and to ship projections to the superior colliculus and the brainstem areas controlling saccades. An anterior system originates in the frontal eye area and tasks, each immediately and through the superior colliculus, to the brainstem saccadic generators. This pathway also passes indirectly through the basal ganglia to the superior colliculus. Projections from the frontal cortex affect cells within the pars reticularis of the substantia nigra, through a relay within the caudate nucleus. An inhibitory pathway from the pars reticulata tasks on to the superior colliculus. This may be a gating circuit related to voluntary saccades, particularly of the memory-guided sort. A posterior pathway originates in the posterior eye field and passes to the brainstem saccadic generators through the superior colliculus. The clean pursuit system has developed relatively independently of the saccadic oculomotor system to keep foveation of a shifting target, although there are inevitable interconnections between the two. The first task is to establish and code the velocity and course of a transferring target. This is carried out within the extrastriate visible area generally recognized as the middle temporal visual area (also called visible area V5), which incorporates neurones sensitive to visible goal movement. In humans, this lies immediately posterior to the ascending limb of the inferior temporal sulcus on the occipitotemporal border. The center temporal visible area sends this motion signal to the medial superior temporal visual area, thought to lie superior and a little anterior to the middle temporal visible area within the inferior parietal lobe; damage to this area leads to an impairment of clean pursuit of targets transferring towards the damaged hemisphere. Both the medial superior temporal visible space and the frontal eye subject send direct projections to a bunch of nuclei that lie in the basal a part of the pons. In monkeys, the dorsolateral and lateral teams of pontine nuclei receive direct cortical inputs related to easy pursuit. These nuclei switch the pursuit signal bilaterally to the posterior vermis, contralateral flocculus and fastigial nuclei of the cerebellum. The pursuit sign ultimately passes from the cerebellum to the brainstem, particularly to the medial vestibular nucleus and nucleus propositus hypoglossi, and thence to the paramedian pontine reticular formation and probably on to the ocular motor nuclei. This circuitry subsequently involves a double decussation: firstly, at the stage of the midpons (pontocerebellar neurones), and secondly, in the lower pons (vestibulo-abducens neurones). This primarily three-neurone arc consists of major vestibular neurones that project to the vestibular nuclei, secondary neurones that project from these nuclei on to the abducens and oculomotor nuclei, and tertiary neurones that innervate the extraocular muscle tissue. Vestibular neurones responding to head rotation also respond to optokinetic stimuli, which signifies that the neural substrate is more doubtless to include both the visual and vestibular techniques. Other muscular vessels branch from the lacrimal and supraorbital arteries or from the trunk of the ophthalmic artery. About seven quick posterior ciliary arteries cross close to the optic nerve to attain the eyeball, the place they divide into 15�20 branches. They reach the eyeball on the tendons of the recti, type a circumcorneal subconjunctival vascular zone, and pierce the sclera near the sclerocorneal junction to finish in the main arterial circle of the iris. Abbreviations: lpca, long posterior ciliary artery; m, muscular arteries; p, pial arteries; spca, brief posterior ciliary arteries. Its terminal branches anastomose on the face and scalp with these of the facial, maxillary and superficial temporal arteries, thereby establishing connections between the exterior and internal carotid arteries.
Its hooked up half breast cancer 10 purchase clomiphene 25mg without prescription, or stalk (petiolus) menopause 1800s order 50mg clomiphene amex, is long and narrow and is related by the elastic thyroepiglottic ligament to the again of the laryngeal prominence of the thyroid cartilage just below the thyroid notch women's health center mount carmel east cheap clomiphene. Its sides are attached to the arytenoid cartilages by aryepiglottic folds (which comprise the aryepiglottic muscle) pregnancy symptoms at 5 weeks buy clomiphene 100 mg on line. Its free higher anterior menstruation every 14 days purchase clomiphene us, or lingual menstrual relief purchase clomiphene pills in toronto, floor is covered by mucosa (the epithelium is nonkeratinized stratified squamous), which is mirrored on to the pharyngeal aspect of the tongue and the lateral pharyngeal partitions as a median glossoepiglottic, and two lateral glossoepiglottic, folds. This arrangement produces a depression, the vallecula, on each side of the median and lateral folds. The lower a part of its anterior floor, behind the hyoid bone and thyrohyoid membrane, is linked to the higher border of the hyoid by an elastic hyoepiglottic ligament, and separated from the thyrohyoid membrane by adipose tissue, which constitutes the clinically essential pre-epiglottic house. The laryngeal cartilages are the midline, single thyroid, cricoid and epiglottic cartilages, and the paired arytenoid, cuneiform, corniculate and tritiate cartilages. The attachments of the false vocal folds (vestibular ligaments; above) and the true vocal folds (vocal ligaments; below) are proven in A, posterior facet. Functions of the epiglottis During swallowing, the hyoid bone strikes upwards and forwards, and the epiglottis is bent posteriorly as a result of passive stress from the bottom of the tongue and energetic contraction of the aryepiglottic muscle tissue. Normally, the epiglottis diverts food and liquids away from the laryngeal inlet and into the lateral meals channels. Ardran and Kemp (1967) argue that, though safe swallowing is possible with out an epiglottis, more care has to be taken whilst eating. Cricoid lamina the cricoid lamina is roughly quadrilateral in define, and 2�3 cm in vertical dimension. It bears a posterior median vertical ridge that creates posterior concavities on both side. The two fasciculi of the longitudinal layer of oesophageal muscle fibres (muscularis externa) are attached by a tendon to the upper part of the ridge. Posterior cricoarytenoid attaches to a shallow melancholy on either facet of the ridge. A discernible circular synovial facet, dealing with posterolaterally, generally marks the junction of the lamina and arch; it signifies the positioning where the cricoid articulates with the inferior thyroid cornu. The posterosuperior aspect of the lamina presents a shallow median notch, on both sides of which is a easy, oval, convex side, directed upwards and laterally, for articulation with the base of an arytenoid cartilage. This projection is most distinct at its upper end, and is nicely marked in males however less seen in women. Above, the laminae are separated by a V-shaped superior thyroid notch or incisure. Posteriorly, the laminae diverge, and their posterior borders are prolonged as slender horns: the superior and inferior cornua. A shallow ridge, the oblique line, curves downwards and forwards on the external floor of every lamina; it runs from the superior thyroid tubercle, lying slightly anterior to the foundation of the superior cornu, to the inferior thyroid tubercle on the inferior border of the lamina. Often, the indirect line is barely seen or is only a faint line and only the tubercles are prominent. The thyroepiglottic ligament, the paired vestibular and vocal ligaments, the thyroarytenoid, thyroepiglottic and vocalis muscle tissue, and the stalk of the epiglottis are all attached to the inside floor of the cartilage, in the angle between the laminae. The inferior border of each lamina is concave posteriorly and nearly straight anteriorly; the two components are separated by the inferior thyroid tubercle. Anteriorly, the thyroid cartilage is connected to the cricoid cartilage by the median (anterior) cricothyroid ligament, which is the thickened medial portion of the conus elasticus. The anterior border of each thyroid lamina fuses with its partner at an angle of approximately 90� in males and roughly 120� in girls. The shallower angle in men is associated with the bigger laryngeal prominence, the higher size of the vocal cords, and the resultant deeper pitch of the voice. The posterior border is thick and rounded, and receives fibres of stylopharyngeus and palatopharyngeus. The superior cornu, which is lengthy and slim, curves upwards, backwards and medially, and ends in a conical apex to which the lateral thyrohyoid ligament is attached. The inferior cornu is brief and thick, and curves down and barely anteromedially. During infancy, some older authors describe a narrow, rhomboidal, flexible strip, the intrathyroid cartilage, lying between the 2 thyroid laminae and joined to them by fibrous tissue. Subglottic stenosis Congenital malformation of the cricoid cartilage could end in severe narrowing of the subglottic airway and respiratory obstruction, which, in severe circumstances, is present from delivery. Acquired subglottic stenosis is more widespread and is the outcomes of trauma and scarring following prolonged endotracheal intubation for the purposes of ventilation of untimely babies (Walner et al 2009). The posterior surface, which is triangular, clean and concave, is roofed by transverse arytenoid. The anterolateral floor is convex and rough, and bears, close to the apex of the cartilage, an elevation from which a crest curves back, down and then forwards to the vocal course of. The upper is triangular (fovea triangularis), and the vestibular ligament is attached to it. The lower is rectangular (fovea oblonga), and vocalis and lateral cricoarytenoid are connected to it. The medial floor is slim, easy and flat, and is roofed by mucosa; its decrease edge forms the lateral boundary of the intercartilaginous a part of the rima glottidis. The base is concave, with a smooth surface for articulation with the lateral a half of the higher border of the cricoid lamina. Its spherical, outstanding lateral angle, or muscular course of, tasks backwards and laterally; it offers attachment to posterior cricoarytenoid behind and lateral cricoarytenoid in entrance. The vocal ligament is attached to its pointed anterior angle (vocal process), which projects horizontally forwards. The apex curves backwards and medially, and articulates with the corniculate cartilage. It is smaller, but thicker and stronger, than the thyroid cartilage, and has a slim curved anterior arch and a broad, flatter posterior lamina. They lie within the posterior parts of the aryepiglottic mucosal folds, and are typically fused with the arytenoid cartilages. Cricoid arch the cricoid arch is vertically slender in front (5�7 mm in height) and widens posteriorly towards the lamina. Cricothyroid is hooked up to the exterior facet of its front and sides, and cricopharyngeus (part of the inferior pharyngeal constrictor) is attached behind cricothyroid. The inferior border of the cartilage is type of horizontal and is circular in define, whereas the higher border is extra elliptical. Their capabilities are unknown, although they might serve to strengthen this connection. They are the small, paired sesamoid cartilages, discovered on the lateral edges of the arytenoid cartilages, and a single interarytenoid cartilage, enclosed by the cricopharyngeal ligament. Each joint is enclosed by a capsular ligament and strengthened by a ligament that, although historically referred to as the posterior cricoarytenoid ligament, is essentially medial in position. The cricoid aspects are elliptical, convex and obliquely directed laterally, anteriorly and downwards. The first is rotation of the arytenoid cartilages at proper angles to the lengthy axis of the cricoid facet (dorso-medio-cranial to ventrolatero-caudal), which, because of its obliquity, causes every vocal process to swing laterally or medially, thereby growing or decreasing the width of the rima glottidis. This motion is sometimes referred to as a rocking motion of the arytenoid cartilages. There can be a gliding movement, by which the arytenoids method or recede from one another, the direction and slope of their articular surfaces imposing a forward and downward motion on lateral gliding. When viewed from above, foreshortening can give the phantasm that the arytenoid cartilages are rotating about their vertical axes, however the form of the aspects prevents such motion occurring (Selbie et al 1998). However, some authors preserve that rotatory motion about a vertical axis can happen (Liu et al 2013). The posterior cricoarytenoid ligaments restrict forward movements of the arytenoid cartilages on the cricoid cartilage. Initially, it includes the decrease and posterior a half of the thyroid cartilage, and subsequently spreads to contain the remaining cartilages, calcification of the arytenoid cartilage beginning at its base. The degree and frequency of calcification of the thyroid and cricoid cartilages seem to be less in females. There is some proof to counsel that a predilection for tumour invasion could also be enhanced by calcification of the laryngeal cartilages (Hatley et al 1965). The tip and upper portion of the vocal means of the arytenoid cartilage consists of non-calcifying, elastic cartilage. The major movement on the joint is rotation round a transverse axis that passes transversely via each cricothyroid joints. There is a few controversy as to whether the cricoid or thyroid cartilage rotates extra. When the joint is in a neutral place, the ligaments are slack and the cricoid can glide, to a restricted extent, in horizontal and vertical directions on the thyroid cornua. The impact of these movements is to lengthen the vocal folds, supplied the arytenoid cartilages are stabilized on the cricoarytenoid joint. The capsules of the laryngeal joints comprise quite a few lamellated (Pacinian) corpuscles, Ruffini corpuscles and free nerve endings (Bradley 2000). It can additionally be interconnected by intrinsic ligaments and fibroelastic membranes, of which the thyrohyoid and quadrangular membranes, along with the conus elasticus, are the most vital. The form of the facet determines the diploma of horizontal and vertical gliding potential at the joint, and thus the degree of elongation of the vocal folds in every case, which has been shown to differ between 3% and 12%. It has been instructed that this variation could be one factor in determining the success of cricothyroid approximation surgical procedure geared toward elevating the pitch of the voice in male to feminine transsexuals (Chen et al (2012), Hammer et al (2010), Windisch et al (2010), Storck et al (2011)). The named ligaments are the median (anterior) cricothyroid ligament, the hyoepiglottic and thyroepiglottic ligaments, and the cricotracheal ligament. It thus ascends behind the concave posterior floor of the hyoid, separated from its physique by a bursa that facilitates the ascent of the larynx during swallowing. Its inner floor is related to the lingual floor of the epiglottis and the piriform fossae of the pharynx. The spherical, cord-like, elastic lateral thyrohyoid ligaments type the posterior borders of the thyrohyoid membrane, and connect the ideas of the superior thyroid cornua to the posterior ends of the higher hyoid cornua. It forms a discontinuous sheet, separated on each side of the larynx by a horizontal cleft between the vestibular and vocal ligaments. Its higher half, the quadrangular membrane, lies within the walls of the higher a part of the laryngeal cavity, the laryngeal vestibule, and extends between the arytenoid cartilages and the edges of the epiglottis. Its lower half, the conus elasticus, lies within the walls of the decrease part of the laryngeal cavity, the infraglottic cavity, and connects the thyroid, cricoid and arytenoid cartilages. C Hyoid bone, higher cornu Thyrohyoid membrane Opening for neurovascular bundle Quadrangular membrane Corniculate cartilage Muscular means of arytenoid cartilage Vocal means of arytenoid cartilage Cricotracheal ligament Quadrangular membrane Each quadrangular membrane passes from the lateral margin of the epiglottis to the apex and fovea triangularis of the ipsilateral arytenoid cartilage. The upper border slopes posteriorly to type the aryepiglottic ligament, which constitutes the central element of the aryepiglottic fold. Posteriorly, it passes via the fascial plane of the oesophageal suspensory ligament, and helps to type the median corniculopharyngeal ligament, which extends into the submucosa adjacent to the cricoid cartilage. The decrease border of the quadrangular membrane forms the vestibular ligament within the vestibular fold. Cut surface of hyoid bone Epiglottis Thyroepiglottic ligament Cut floor of thyroid cartilage Vestibular ligament Vocal ligament Conus elasticus Cricoid cartilage Cricothyroid membrane and conus elasticus the conus elasticus is that part of the fibroelastic membrane discovered within the lower a part of the cavity of the larynx. The terminology used to describe this construction is complicated, as completely different phrases could additionally be used to describe apparently similar structures. Two synonyms for the conus elasticus which might be generally discovered in the literature are the cricovocal membrane and the cricothyroid membrane. The time period conus elasticus is regularly applied to the lateral elements, while the median half is often known as both the median or the anterior cricothyroid ligament. The supraglottis is a clinical time period typically used in tumour staging; it refers to all those elements of the larynx that lie above the glottis and thus comprises the laryngeal inlet (formed of the laryngeal floor of the epiglottis and arytenoid cartilages, and the laryngeal aspects of the aryepiglottic folds), the laryngeal vestibule and the vestibular folds. Other phrases which might be used clinically in tumour staging are the glottis, outlined as the anterior and inferior surfaces of the true vocal folds and the anterior and posterior commissures; and the subglottis, defined because the region under the glottis that extends to the inferior border of the cricoid cartilage. Median (anterior) cricothyroid ligament the median or anterior cricothyroid ligament is the thickened central portion of the conus elasticus. The partitions of the cavity are fashioned of the fibroelastic membranes described above and lined with mucous membrane that folds over the free edges of these membranes inside the larynx. On both facet, the continuity of the fibroelastic membrane is interrupted between the higher vestibular and lower true vocal folds. The folds project into the lumen of the cavity and divide it into higher and lower parts, separated by a middle portion between the two units of folds that leads into the laryngeal ventricle. The higher folds are the vestibular (ventricular or false vocal) folds; the median aperture between them is the rima vestibuli. The decrease pair are the (true) vocal folds (or vocal cords), and the fissure between them is the rima glottidis or glottis. Laryngeal ventricle Laryngeal inlet (aditus) the upper part of the laryngeal cavity is entered by the laryngeal inlet (aditus laryngis), an roughly triangular aperture between the larynx and pharynx. This faces backwards and considerably upwards because the anterior wall of the larynx is much longer than the posterior (and slopes downwards and forwards in its upper half because of the oblique inclination of the epiglottis). The inlet is bounded anteriorly by the higher fringe of the epiglottis, posteriorly by the transverse mucosal fold between the 2 arytenoids (posterior commissure), and on each side by the edge of a mucosal ridge, the aryepiglottic fold, that runs between the aspect of the epiglottis and the apex of the arytenoid cartilage. The midline groove between the two corniculate tubercles is termed the interarytenoid notch. Inferiorly, the conus elasticus attaches to the superior cricoid arch and the cricoid lamina. Between these anterior and posterior attachments, the upper edges of the conus elasticus are free, thickened and aligned horizontally, forming the vocal ligaments. Each vocal ligament lies within a mucosacovered vocal fold, coated on its inner floor by the mucosal lining of the larynx, and externally by the lateral cricoarytenoid and thyroarytenoid muscles (Reidenbach 1995). It is loosely adherent to the anterior floor of the epiglottis but firmly hooked up to its anterior surface and the floor of the valleculae.
The foramen may be in the occipital or occipitotemporal suture; it might generally be parasutural or may be absent women's health clinic joondalup discount clomiphene 50mg amex. The mastoid part initiatives down because the conical mastoid process and is bigger in adult males menstrual extraction clomiphene 50mg. Sternocleidomastoid menstrual 10 days discount 100 mg clomiphene free shipping, splenius capitis and longissimus capitis are all hooked up to its lateral floor menstrual like cramps after hysterectomy cheap clomiphene 25mg, and the posterior belly of digastric is attached to a deep mastoid notch on its medial aspect menstruation and breastfeeding clomiphene 25 mg online. The occipital artery runs in a shallow occipital groove that lies medial to the mastoid notch pregnancy joint pain cheap 50 mg clomiphene mastercard. The inside floor of the mastoid course of bears a deep, curved sigmoid sulcus for the sigmoid venous sinus; the sulcus is separated from the underlying innermost mastoid air cells by a skinny lamina of bone. The posterior border can additionally be serrated and articulates with the inferior border of the occipital bone between its lateral angle and jugular course of. The mastoid factor is fused with the descending strategy of the squamous half; below, it seems within the posterior wall of the tympanic cavity. The posterior border, intermediate in size, bears a sulcus medially, which types, together with the occipital bone, a gutter for the inferior petrosal sinus. Behind this, the jugular fossa contributes (together with the occipital bone) to the jugular foramen and is notched by the glossopharyngeal nerve. Bone on both or both sides of the jugular notch might meet the occipital bone and divide the jugular foramen into two or three elements. The anterior border is joined laterally to the squamous a half of the temporal bone on the petrosquamosal suture; medially, it articulates with the greater wing of the sphenoid bone. Two canals exist at the junction of the petrous and squamous elements, one above the opposite, separated by a thin osseous plate and both resulting in the tympanic cavity; the upper canal contains tensor tympani, whereas the decrease canal is the pharyngotympanic tube. It is inclined superiorly and anteromedially, and has a base, apex, three surfaces (anterior, posterior and inferior) and three borders (superior, posterior and anterior). The base would correspond to the part that lies on the base of the skull and is separated from the squamous half by a suture. The subsequent development of the mastoid processes signifies that the exact boundaries of the bottom are now not identifiable. The apex, blunt and irregular, is angled between the posterior border of the greater wing of the sphenoid and the basilar a part of the occipital bone. It accommodates the anterior opening of the carotid canal and limits the foramen lacerum posterolaterally. Bone anterolateral to this impression roofs the anterior a half of the carotid canal however is often poor. A ridge separates the trigeminal impression from one other hollow behind, which partly roofs the internal acoustic meatus and cochlea. Laterally, the anterior surface roofs the vestibule and, partly, the facial canal. Between the squamous half laterally and the arcuate eminence and the hollows simply described medially, the anterior surface is shaped by the tegmen tympani, a thin plate of bone that forms the roof of the mastoid antrum, and extends forwards above the tympanic cavity and the canal for tensor tympani. The lateral margin of the tegmen tympani meets the squamous half at the petrosquamosal suture, turning down in front because the lateral wall of the canal for tensor tympani and the osseous a half of the pharyngotympanic tube; its lower edge is within the squamotympanic fissure. Anteriorly, the tegmen bears a narrow groove related to the higher petrosal nerve (which passes posterolaterally to enter the bone by a hiatus anterior to the arcuate eminence). The posterior slope of the arcuate eminence overlies the posterior and lateral semicircular canals. Lateral to the eminence, the posterior part of the tegmen tympani roofs the mastoid antrum. The posterior surface contributes to the anterior a half of the posterior cranial fossa and is steady with the internal surface of the mastoid half. A small slit leading to the vestibular aqueduct lies behind the opening of the meatus, nearly hidden by a thin plate of bone. This incorporates the saccus and ductus endolymphaticus, together with a small artery and vein. The terminal half of the saccus endolymphaticus protrudes via the slit between the periosteum and dura mater. Near the apex of the petrous part, a quadrilateral area is partly related to the attachment of levator veli palatini and the cartilaginous pharyngotympanic tube, and partly related to the basilar a part of the occipital bone by dense fibrocartilage. Behind this region is the large, circular opening of the carotid canal, and behind the opening of the canal is the jugular fossa, which is of variable depth and dimension, and incorporates the superior jugular bulb. The inferior ganglion of the glossopharyngeal nerve lies in a triangular despair anteromedial to the jugular fossa (below the internal acoustic meatus). At its apex is a small opening into the cochlear canaliculus, occupied by the perilymphatic duct (a tube of dura mater) and a vein draining from the cochlea to the interior jugular vein. A canaliculus for the tympanic nerve from the glossopharyngeal nerve lies on the ridge between the carotid canal and the jugular fossa. The mastoid canaliculus for the auricular branch of the vagus nerve is laterally positioned within the jugular fossa. Behind the jugular fossa, the tough quadrilateral jugular surface is roofed by cartilage that joins it to the jugular means of the occipital bone. Tympanic part the tympanic part of the temporal bone is a curved plate below the squamous part and anterior to the mastoid process. Behind, it fuses with the squamous half and mastoid course of, and is the anterior limit of the tympanomastoid fissure. Its concave posterior floor forms the anterior wall, flooring and part of the posterior wall of the exterior acoustic meatus. The tympanic membrane is hooked up to a slender tympanic sulcus on its medial floor. The quadrilateral concave anterior floor is the posterior wall of the mandibular fossa and should contact the parotid gland. Its rough lateral border forms most of the margin of the osseous a half of the exterior acoustic meatus and is continuous with its cartilaginous half. Laterally, the upper border is fused with the again of the postglenoid tubercle; medially, it forms the posterior fringe of the petrotympanic fissure. The inferior border is sharp and splits laterally to form, at its root, the sheath of the styloid course of (vaginal process). The stylomastoid foramen lies between the styloid and mastoid processes; it represents the exterior finish of the facial canal, incorporates a half of the aponeurosis of the posterior stomach of digastric, and transmits the facial nerve and stylomastoid artery. Often virtually straight, it can show a curvature, an anteromedial concavity being commonest. Its proximal half (tympanohyal) is ensheathed by the tympanic plate, particularly anterolaterally, whereas muscles and ligaments are attached to its distal part (stylohyal). The styloid course of is roofed laterally by the parotid gland; the facial nerve crosses its base; the external carotid artery crosses its tip, embedded in the parotid; and medially, the method is separated from the start of the interior jugular vein by the attachment of stylopharyngeus. External acoustic meatus the temporal bone accommodates the bony (osseous) a half of the exterior acoustic meatus. The squamous half is ossified in a sheet of condensed mesenchyme from a single centre near the zygomatic roots, which seems within the seventh or eighth week in utero. The petromastoid part has a number of centres that appear in the cartilaginous otic capsule through the fifth month; as many as 14 have been described. Key: 1, helix; 2, crus of helix; three, auricular tubercle; 4, antihelix; 5, crura of antihelix; 6, triangular fossa; 7, scaphoid fossa; 8, concha of auricle; 9, exterior acoustic meatus; 10, tragus; 11, antitragus; 12, intertragic notch; 13, lobule of auricle. The styloid course of develops from two centres at the cranial end of cartilage within the second visceral or hyoid arch; a proximal centre for the tympanohyal appears earlier than delivery, and another, for the distal stylohyal, appears after delivery. The tympanic ring unites with the squamous part shortly before start, and the petromastoid fuses with it and the tympanohyal through the first 12 months. Once ossified, the tympanic cavity, mastoid antrum and the posterior finish of the pharyngotympanic tube turn into surrounded by bone. The petrous half types the roof, flooring and medial wall of the cavity, while the squamous and tympanic parts, together with the tympanic membrane, form its lateral wall. At start, the middle and inner ears are grownup measurement, and the tympanic cavity, mastoid antrum, tympanic membrane and auditory ossicles are all almost adult measurement. The inside acoustic meatus is approximately 6 mm in horizontal diameter, four mm in vertical diameter and 7 mm in size at start, and the grownup diameters are 7. After delivery and apart from general growth, the tympanic ring extends posterolaterally to turn out to be cylindrical, rising into a fibrocartilaginous tympanic plate, which varieties the adjacent a half of the external acoustic meatus at this stage. Thus, for a time, a gap (foramen of Huschke) exists in the ground; it usually closes at about the fifth yr but is usually everlasting. The tympanic plate ensheathes the styloid process by posterior extension, and extends medially over the petrous bone to the carotid canal. Initially, the mandibular fossa is shallow, dealing with extra laterally, but it then deepens and finally faces downwards. Posteroinferiorly, the squamous half grows down behind the tympanic ring to type the lateral wall of the mastoid antrum. The mastoid half is at first flat, so that the stylomastoid foramen and rudimentary styloid process are immediately behind the tympanic ring. The lateral mastoid area grows downwards and forwards to kind the mastoid course of, which means that the styloid process and stylomastoid foramen become inferior. In the neonate, the petrous and squamous components of the temporal bone are normally partially separated by the petrosquamous fissure, which opens instantly into the mastoid antrum of the middle ear. There is a bimodal development pattern within the lateral floor dimensions of the temporal bone in youngsters; marked will increase in dimension occur from birth to the age of 4 years but little growth is seen between the ages of four and 20 years (Simms and Neely 1989). The neonatal internal acoustic meatus is about half the length of its grownup counterpart. The course of the pharyngotympanic tube is horizontal within the newborn, whereas in the adult it passes from the center ear downwards, forwards and medially. The auricle tasks to a variable and particular person diploma from the side of the head. It collects sound waves and conducts them alongside the external acoustic meatus inwards in course of the ear drum, the tympanic membrane. The auricle thus acts as the primary element of a collection of stimulus modifiers within the auditory equipment. The antihelix is a curved prominence, parallel and anterior to the posterior a part of the helix; it divides above into two crura, which flank a depressed triangular fossa. The antihelix encircles the deep, capacious concha of the auricle, which is incompletely divided by the crus or anterior end of the helix. The conchal area above this, the cymba conchae, overlies the suprameatal triangle of the temporal bone, which could be felt through it, and which overlies the mastoid antrum. The tragus is a small curved flap beneath the crus of the helix and in front of the concha; it projects posteriorly, partly overlapping the meatal orifice. The antitragus is a small tubercle reverse the tragus and is separated from it by the intertragic incisure or notch. It is delicate, unlike the majority of the auricle, which is supported by elastic cartilage and is agency. At birth, many of the linear dimensions of the auricle are roughly three-quarters of their grownup size; the length and top of the tragus are less than half of their grownup measurement. Width dimensions mature between the ages of 5 and eleven years; length dimensions mature between 12 and sixteen years (Purkait 2013). Common congenital anomalies Developmental anomalies of the branchial arches may produce a grossly misshapen or microtic auricle, sometimes with associated anomalies of the middle ear and vital listening to loss. A variety of common anomalies have been acknowledged; they carry descriptive names or eponyms (Porter and Tan 2005) Table 37. The cranial side of the cartilage bears the eminentia conchae and eminentia scaphae, which correspond to the depressions on the lateral floor. The two eminences are separated by a transverse furrow, the sulcus antihelicis transversus, which corresponds to the inferior crus of the antihelix on the lateral floor. The eminentia conchae is crossed by an indirect ridge, the ponticulus, for the attachment of auricularis posterior. There are two fissures within the auricular cartilage, one behind the crus of the helix and one other in the tragus. Skin the skin of the auricle continues into the exterior acoustic meatus to cowl the outer surface of the tympanic membrane. It is thin, has no dermal papillae, and is closely adherent to the cartilaginous and osseous parts of the canal; irritation of the canal pores and skin could be very painful because of this attachment to the underlying buildings. The thick subcutaneous tissue of the cartilaginous a half of the meatus accommodates quite a few ceruminous glands that secrete wax, or cerumen. Ducts open either on to the epithelial floor or into the nearby sebaceous gland of a hair follicle. Antibacterial properties have been attributed to cerumen however the evidence for that is missing (Campos et al 2000, Pata et al 2003). Dry wax is frequent in East Asians, whereas the moist sort is extra widespread in other ethnic teams (Yoshiura et al 2006). Overproduction, accumulation or impaction of wax could utterly occlude the meatus. This might hinder sound from reaching the tympanic membrane and so limit its pure vibration. Although ceruminous glands and hair follicles are largely restricted to the cartilaginous meatus, a few small glands and nice hairs are also current within the roof of the lateral a part of the bony meatus. The warm, humid environment of the relatively enclosed meatal air aids the mechanical responses of the tympanic membrane. Ligaments Anterior and posterior extrinsic ligaments join the auricle with the temporal bone. The anterior ligament extends from the tragus and the backbone of the helix to the basis of the zygomatic means of the temporal bone. The posterior ligament passes from the posterior surface of the concha to the lateral floor of the mastoid process. Two main intrinsic ligaments join particular person auricular cartilages: a powerful fibrous band passes from the tragus to the helix, thereby completing the meatus anteriorly and forming a half of the boundary of the concha; and one other band passes between the antihelix and the tail of the helix. It is linked to the encircling elements by ligaments and muscle tissue, and is steady with the cartilage of the exterior acoustic meatus.
The inspecting finger identifies first the subclavian pulse after which the nerves 32 menstrual cycle clomiphene 100 mg sale, while rolling the fingertip laterally (see Video 46 womens health 15 minute workout buy 100mg clomiphene free shipping. Divisions of the brachial plexus the trunks divide into anterior and posterior divisions menstrual gas cramps 100 mg clomiphene overnight delivery. The higher trunk divides 2�3 cm above the clavicle women's health boca raton buy clomiphene amex, and the divisions of the center and lower trunk are fashioned deep to the clavicle xenoestrogens menopause purchase 50mg clomiphene fast delivery. It passes posteriorly menstrual tissue discharge cheap clomiphene 50mg otc, pier cing scalenus medius, to run down in the plane between levator scapu lae and serratus posterior superior and the posterior scalene muscle tissue. The dorsal scapular nerve inside vates the rhomboid muscular tissues and, along with branches from C3 and C4, it provides levator scapulae. BloodsupplyThe blood supply of the brachial plexus is derived from vessels arising from the subclavian and vertebral arteries. Important branches pass from the vertebral artery to the rami of C5 and C6 and the more proximal cervical nerves. Extensive contributions come from the suprascapular and superficial cervical arteries that come up from the thyrocervical trunk. In a minimum of onethird of cases, the superficial cervical and dorsal scapular arteries come up from the thyrocervical trunk because the transverse cervical artery. In scientific terms, the arteries arising from the thyrocervical trunk have turn into the lifeline to the upper limb and must be preserved during operations in circumstances when the ruptured subclavian artery has not been repaired. The dorsal scapular artery could arise from the third part of the subclavian artery to cross between the upper and center trunks of the brachial plexus. The rami enter the posterior triangle of the neck between scalenus anterior and medius. Rami from C7 and C8 are the largest (C8 incorporates about 30,000 myeli nated axons) and people from C5 and T1 are the smallest (between 15,000 and 20,000 myelinated axons). An grownup brachial plexus con tains between 120,000 and one hundred fifty,000 myelinated axons, of which 25% innervate the shoulder girdle and glenohumeral joint. The proportion of motor fibres is biggest in C5 and C8; the sensory contribution is biggest in C7. A complex interchange of branches, earlier than the principle nerves of the higher limb are fashioned, produces the trunks, divisions and cords of the brachial plexus. Note the sequence: ventral (anterior) main rami; trunks; divisions; cords; nerves. The trunks are higher, middle and decrease, and the cords are lateral, medial and posterior, in accordance with their position in relation to the axillary artery, which is variable. The posterior division of the lower trunk is persistently smaller than the anterior division and is absent in about 10% of circumstances. Cords of the brachial plexus 782 the cords are fashioned by the confluence of divisions: the lateral twine from the anterior divisions of the higher and middle trunks; the pos terior twine by all three posterior divisions; and the medial wire by the anterior division of the decrease trunk and, generally, by a branch from the anterior division of the center trunks. The divisions of the trunks and the formation of the cords characterize an necessary anatomical and practical differentiation. The posterior divisions and posterior wire innervate postaxial (extensor) musculature; the anterior divisions and the lateral and medial cords innervate preaxial (flexor) musculature. The formation and relations of the three cords are variable and, certainly, their designations considerably misleading. Immediately inferior to the clavicle, the posterior wire is lateral, the medial twine is posterior, and the lateral wire is anterior, in relation to the axillary artery; the cords assume their applicable relations concerning the axillary artery deep to pectoralis minor. There is appreciable variation in this association; mostly, the axillary artery lies anterior to the three cords and the median nerve. The branches of the posterior cord, the biggest of the three trunks, are constant. The branches of the medial cord are normally predictable; the medial pectoral nerve and medial cutaneous nerve of the forearm are succeeded by the division into the medial root of the median nerve and the ulnar nerve. The greatest variation in for mation of trunk nerves is discovered inside the lateral cord. Occasionally, the musculocutaneous nerve arises extra distally than ordinary, springing either directly from the lateral wire as two or three branches or even from the median nerve itself. Sometimes, the highest of these branches enters coracobrachialis no extra than 2 or three cm beneath the coracoid course of. The lateral root of the median nerve could come up as two or three branches, and in some circumstances, it appears as a branch of the musculo cutaneous nerve. The first 8�10 cm of the nerve may not have a nutrient artery and can, therefore, be relatively avascular if the nerve is transected at its origin. The nerve lies closest to the bone the place it pierces the lateral intermuscular septum to pass through a short tunnel bounded by bone and unyielding fascia. The nerve to the medial head of triceps arises within the axilla and accompanies the main nerve because it passes via the intermuscular septum. The nerve(s) to the lateral head of triceps arise(s) throughout the spiral groove; that to the lateral head just earlier than the radial nerve reenters the anterior compartment. They innervate the pores and skin of the lower lateral arm and the posterior side of the forearm. These branches spring from the principle nerve as it runs within the valley between brachialis anteriorly and brachioradialis posteriorly, which is a web site where the nerve could also be palpated. Variation exists within the cutaneous innervation of the dorsal aspects of the digits. Here, the radial nerve is proven to provide all five digits; the pores and skin of the dorsum of the ring and little fingers is frequently innervated by the dorsal department of the ulnar nerve. The superficial radial nerve innervates the skin over the dorsum of the radial half of the ring, center and index fingers, and that of the thumb so far as the distal interphalan geal joint. It usually offers important sensation on the skin of the thumb web space and the skin on the adjoining sides of the thumb and index. It innervates teres minor and deltoid, and the skin overlying deltoid on the lateral side of the shoulder. The major nerve contin ues in the aircraft between the superficial and deep digital flexors, sup plying two or three branches to the previous. The palmar cutaneous nerve arises about three cm proximal to the proximal wrist crease, and passes lateral to the principle nerve and superficial to the flexor retinaculum to innervate the skin of the proximal palm. The median nerve passes deep to the flexor retinaculum, into the carpal tunnel, to enter the palm. The nerve to the thenar muscular tissues arises inside, or simply distal to , the tunnel, often on the lateral aspect of the primary nerve. It is palpable on the flexor facet of the elbow, deep to the bicipital aponeurosis, and is accompanied by the brachial artery on this section of the limb. The median nerve is palpable at the wrist where it emerges from behind the superficial flexor tendons, just lateral to palmaris longus. It inclines posteromedially to perforate the medial inter muscular septum about 10 cm proximal to the medial epicondyle and passes by way of a fibrous canal, the arcade of Struthers, which is between 5 and 6 cm long. The partitions of the canal embody the medial intermus cular septum and the fascial sheath investing the medial head of triceps brachii. The nerve next passes behind the medial epicondyle, then between the capsule of the humeroulnar joint and the overlying arcuate ligament, which joins the 2 origins of flexor carpi ulnaris, i. It is accompanied by the supe rior collateral ulnar vessels within the decrease third of the arm as far as the doorway to the cubital tunnel, after which by the posteroinferior ulnar collateral vessels. The anastomosis between these vessels permits the usage of many of the ulnar nerve as a free vascularized graft, in circumstances where the father or mother nerves, C7, C8 and T1, are irreparable. In the forearm, the nerve runs with the ulnar vessels in a welldefined sheath deep to the anterior margin of flexor carpi ulnaris. The superficial division innervates the skin of the medial two digits and palmaris brevis. The deep branch passes, with the deep department of the ulnar artery, between abductor and flexor digiti minimi, pierces opponens digiti minimi after which runs across the deep palmar house with the deep palmar arch. It innervates all of the muscular tissues of the hand, apart from the first and second (index and middle) lumbricals, abductor pollicis brevis, opponens pollicis and a half of flexor pollicis brevis. The lateral root conveys a lot of the sympathetic fibres to the median dis tribution within the hand. The medial root carries fibres to the skin of the medial side of the middle and the lateral side of the ring finger, and also fibres to palmaris longus, flexor digitorum superficialis and the lateral a half of flexor digitorum profundus, flexor pollicis longus, prona tor quadratus and the median innervated muscle tissue inside the hand. The first branch of the median nerve, to pronator teres, arises 2�3 cm above the medial epicondyle. A second department, or leash of branches, arises at the stage of the tip of the medial epicondyle and innervates palmaris longus, flexor carpi radialis and flexor digitorum superficialis. Just proxi mal to the tendinous arcade of flexor digitorum superficialis, the median nerve provides off the anterior interosseous nerve. A department to the medial head of triceps is commonly given off in the distal third of the arm. There are often two neurovascular pedicles to flexor carpi ulnaris, the primary on the exit from the cubital tunnel, the second about onequarter of the way in which down the forearm. The dorsal cutaneous nerve arises about 6 cm proximal to the ulnar styloid, winds around the ulna, at first deep to flexor carpi ulnaris, and then subcutaneously. It innervates the skin of the dorsum of the hand and little finger, and the medial aspect of the ring finger. There is a crucial relation right here: a posterior branch of the medial cutaneous nerve of the forearm sweeps throughout the cubital tunnel and gets in the means in which of medial surgical exposures. The ulnar nerve is palpable at the wrist where it emerges from beneath flexor carpi ulnaris. The dorsal cutaneous department can be palpated in those of slender physique where it crosses the subcutaneous ulna. This division normally happens at the degree of the third a half of axillary artery, the place the lateral wire is lateral to the vessel, however it could occur more proximally, the place the wire is anterior to the second part of the artery. The nerve may also come up as two or three branches from the wire, or even from the median nerve. It supplies coracobrachialis, each heads of biceps brachii and most of brachialis, and continues as the lateral cutaneous nerve of forearm, deep to biceps, before rising lateral to it about eleven cm above the lateral epicondyle, and then working down the lateral aspect of the forearm. The nerve generally innervates the pores and skin over the first metacarpal and the lateral a part of the thenar eminence. The most proximal muscle tissue are supplied by branches of the uppermost rami, and probably the most distal muscle tissue are equipped by branches derived from C8 and T1. The segmental sample of innervation is shown more clearly in the cutaneous supply. C6 usually innervates biceps brachii, brachialis, lateral head of triceps, brachioradialis, extensor carpi radialis longus, and generally contributes to the innervation of pronator teres, supinator and flexor carpi radialis. C7 offers widespread innervation all through the limb; in these unusual situations the place it alone is broken, the patient will notice a rather diffuse loss of operate all through the upper limb, often with out full anaesthesia and sometimes without paralysis of any single important muscle group. C8 innervates the extensors of the digits and of the thumb in at least onethird of instances. Muscles, movements and segmental innervation within the upper limb are set out in Tables 46. Autonomicnerves C3 C4 C5 T2 T3 T4 T2 T5 T6 T7 T8 C6 T1 T9 T10 T11 C6 T12 L1 the preganglionic sympathetic influx to the upper limb is derived from neurones in the intermediolateral horn of spinal wire segments T2�6/7. Myelinated preganglionic axons cross from the spinal nerves to the thoracic sympathetic chain through white rami communicates. They move up the chain to the center cervical and inferior cervical (stellate or cervico thoracic) ganglia, the place they relay. The unmyelinated postganglionic fibres are distributed by gray rami communicantes to the nerves of origin of the brachial plexus. The sympathetic outflow to the upper limb passes with the peripheral nerves and is especially rich within the palmar skin of the hand and in the skin of the axilla. The acromioclavicular joint line is palpable as an anteroposteriorly aligned ridge or groove. Posteriorly, the palpable acromial angle marks the junction of the acromion with the crest of the scapular spine. From this point, the spine of the scapula, which is subcutaneous and simply visible in skinny topics, is palpable to the medial (vertebral) border of the scapula, the place it lies reverse the backbone of the third thoracic vertebra. The medial border of the scapula, superior to the extent of the scapu lar spine, is covered by trapezius and, subsequently, troublesome to palpate. Although lined by teres major and latissimus dorsi, the inferior angle is palpable when approached inferiorly, and could be seen and/or palpated because it strikes across the thoracic wall during shoulder move ments. Spinal roots have been given the same colour (light peach, dark peach, red) when they innervate a muscle to an identical extent. In the acute injury, the item of the clinician should be to acknowledge the very fact of injury as soon as potential after the event, and later, to go on to decide the nerve or nerves affected, the level(s) of harm, and the extent and depth of the lesion(s). The history is important: excessive injury transfer, open fracture and wounding (be it unintentional, legal, surgi cal, or all three) indicate a severe lesion and recommendation from witnesses or emergency paramedical staff is all the time priceless. Injuries which are poten tially life or limbthreatening complicate closed traction lesions of the supraclavicular brachial plexus in about 20% of cases. It is necessary to seek occult injuries to the head, backbone, thorax, abdomen and pelvis before embarking on remedy of the nerve lesion(s). The early signs of acute nerve damage include: irregular spon taneous sensations; alteration or lack of sensibility; weak point and par alysis; impairment of perform; an awareness of warming and dryness of all or a half of an extremity (sometimes); and pain (sometimes). It can be distinguished from the ache of fracture or dislocation by lack of sensation; painful, spon taneous sensory signs, expressed all through the territory of the nerve; and by lancinating or capturing ache, irradiating into the distribu tion of the nerve. Constant crushing, bursting or burning pain in an in any other case undamaged hand indicates critical and continuing injury to main trunk nerves extra proximally. Progression of sensory loss, with a deep bursting or crushing pain within the muscular tissues of the limb, signi fies critical ischaemia. The major nerves are stretched, ruptured or avulsed from muscle by violent hyperextension�abduction injuries of the arm. The axillary or brachial artery is ruptured in no much less than onethird of fractures of the proxi mal humerus or fracture dislocations of the shoulder. The axillary nerve and posterior circumflex vessels, which move virtually horizontally pos terior to the neck of the humerus, are significantly in danger from anterior dislocation of the pinnacle of the humerus.
Discount clomiphene 25 mg with visa. Upstate Womens Health Network Holiday Message.